

Permanent-high-bed technology to improve intensive vegetable production in tropical lowlands - Effect on water stress, root distribution, and yield
Kleinhenz, V.; Schnitzler, W. H.; Midmore, D. J., 1996
Horticultural Science, 28, 90-98
HORTICULTURAL SCIENCE - KERTÉSZETI TUDOMÁNY 1996. 28. (1–2): 90-98
Permanent-high-bed technology to improve intensive vegetable production in tropical lowlands - Effect on water stress, root distribution, and yield
Kleinhenz V.1, Schnitzler W. H.2 and Midmore D. J.3
1 Asian Vegetable Research and Development Center, PO Box 42, Shanhua, Tainan, Taiwan - 741, R O C
Current address: Institute for Vegetable Science, Technical University Munich, D-85350 Freising, Germany
2 Institute for Vegetable Science, Technical University Munich, D-85350 Freising, Germany
3 Asian Vegetable Research and Development Center, PO Box 42, Shanhua, Tainan, Taiwan - 741, R O C
Current address: Department of Biology, Central Queensland University, Rockhampton, Qld 4702, Australia
Key words: Southeast Asia, rice, cultivation system, soil moisture tension, root length distribution.
Abstract: The effect of permanent high beds between continuously flooded furrows on soil moisture stress, root growth, and yield was compared to traditional flat-bed cultivation in four vegetable crops during 1994/95 at the Asian Vegetable Research and Development Center (AVRDC) in the tropical lowland of Taiwan. A method to integrate soil moisture tension over time and soil depth is presented which, in contrast to earlier attempts, also accounts for stress following overwet soil conditions as they frequently occur in tropical lowlands. The approach of calculating soil moisture stress as an integral of positive and negative deviates from an estimated crop-specific ‘optimum soil moisture tension’ was a suitable tool to explain variation in vegetable yield. The relationship between yield and soil moisture stress was found to be nearly linear for the crops grown during the dry season (vegetable soybean and carrot), but exponential for crops during the rainy season (Chinese cabbage and chili). This hints at the exaggerated effect of extremely low values of soil moisture tension on crop growth. Root growth of all species on flat beds was typically restricted to the topmost soil. Therefore, inclusion of soil moisture tension measurements at greater soil depth (45 cm) did not improve the estimation of soil moisture stress based on the shallow depth (15 cm). In high beds, vegetables developed more profuse root systems than in flat beds. This resulted in better yields of vegetables grown during the rainy season. Better hydraulic properties of high beds in the rainy season have their roots in horizontal removal of excessive soil water when vertical flow is negligible. Gradients of soil moisture tension revealed that the furrows between them acted as a sink for excessive soil water when heavy rainfall occurred and the soil was saturated throughout the profile. During dry periods and when soil moisture was low, the flooded furrows supplied the beds with water. These effects weakened towards the center which was rapidly inundated under wet conditions and more drought prone under dry conditions. Therefore, wider high beds are only advantageous when prolonged times of rainfall or drought do usually not occur.
Introduction
Intensive vegetable production under tropical lowland conditions depends to a large extent on successful management of soil water. Deficient and excessive moisture conditions induce water stress in vegetables and can proceed in close alternation particularly during the rainy summer season when satisfactory production remains beyond the means of most farmers (Midmore et al., 1995). Improved field management practices such as permanent-high-bed technology are one way of increasing vegetable production as shown in some peri-urban lowland production zones in Southeast Asia (Sjahri, 1975; Plucknett and Beemer 1981; Chiu, 1987; Kieft, 1994).
To gauge the effect of field management techniques on alleviating water stress in vegetables, a suitable measure of soil moisture stress has to be developed. If soil moisture is to be related to plant response, soil moisture tension measurements are preferred over moisture content measurements since they are a better measure of the availability of soil water to plants (Gardner, 1960). In this thermodynamic terminology, movement of water in soil, its uptake by plants, and its loss to the atmosphere through transpiration is explained as a change of state from higher to lower free energy (Brady, 1990). Therefore, water moves from lower tension to higher tension (Cassell and Klute, 1986). Increased stomatal resistance during drought periods causes a reduction in carbon dioxide uptake and photosynthesis (Millar and Gardner, 1972), and a reduction in transpiration and nutrient uptake (Hanks, 1991). Therefore, plant growth and crop yield can be constrained by increased soil water tension (Gingrich and Russel, 1957; Rawitz and Hillel, 1969).
Integration of the variability in the soil-plant system over time during the growing period of a crop provides a way to evaluate the effect of soil physical conditions on crop growth (Callebaut et al., 1982). Wadleigh (1946) argued that since soil water tension cannot be maintained constant within the range of available water content, water stress in plants must depend upon the rate of change of soil moisture stress over the growth period. He defined a moisture stress value as an integral of soil moisture tension for time. Taylor (1952 a and b) calculated ‘mean integrated soil moisture tension’ as a double integral for both time (growth period) and soil depth and found significant relationships between soil-moisture-stress values and crop yields. Since the reference point of zero stress was set for zero tension, better yields were associated with lower moisture tension values. However, besides sufficient water to satisfy the needs of transpiration and nutrient uptake, plants also require gaseous exchange between root and soil. Because the above-mentioned estimate of soil water stress takes into account only depletion of available soil moisture, it does not account for stresses caused by excessive soil water conditions with their manifold consequences for crop production (Ponnamperuma, 1984). It follows that, in an environment where soil flooded conditions frequently occur, a soil moisture stress index should also include stress caused by excessive soil moisture. For this, the reference point of zero soil moisture stress should be set for more than zero tension, and the integration of soil moisture tension should include the absolute value of the deviates from the optimum.
The distribution of roots with soil depth mainly determines the water uptake patterns of plants (Gardner, 1964) and density distribution of root lengths coincides with root activity (Richter, 1987). Therefore, root density distribution reflects environmental conditions integrated over the time before measurement (Bathke et al., 1992).
The objective of this study was (1) to develop a suitable method to quantify soil moisture stress including stress caused by overwet soil conditions, (2) to determine the influence of traditional flat beds and permanent high beds on root distribution and yield of vegetables year-round, and (3) to study the hydraulic properties of high beds in a tropical lowland.
Material and methods
Site, systems and crop management
From 1992 to 1995, the influence of different cultivation systems on the production of vegetables year-round was studied at the AVRDC research farm in southern Taiwan (23º N latitude). The soil at the experimental site was an alluvial sandy loam (18 % clay, 27 % silt, 55 % sand). Monthly sums of precipitation and evaporation are presented in Fig. 1. Cultivation systems included traditional flat beds (1.5 m wide, 20-25 cm high), which were constructed before each crop and permanent high beds (50 cm high) with different widths (2.0 or 3.0 m) between continuously flooded furrows (2.0 m wide). During 1994/95 four vegetable crops, namely vegetable soybean (Glycine max. L. Merr; cv. ‘AGS 292’, AVRDC; March to May 1994), Chinese cabbage (Brassica pekinensis Lour. Rupr.; cv. ‘ASVEG No. 1’, AVRDC; May to July 1994), chili (Capsicum annuum L.; cv. ‘Hot Beauty’, Known You Seed Co.; July to December 1994), and carrot (Daucus carota L. ssp. sativus Hoffm. Arcang.; cv. ‘Parano’, Nunhems; January to April 1995) were cultivated with distinct crop row distances (paired rows for carrot) from the edge of the beds (40 cm on flat beds; 40 and 80 cm on 2.0-m-wide high beds; 40, 80 and 120 cm on 3.0-m-wide high beds). Aquatic crops cultivated in the flooded furrows were taro (Colocasia esculenta (L.) Schott; December 1993 to July 1994) and rice (Oryza sativa L.; August to December 1994 and February to June 1995). Standing water was maintained at a height of a few centimeters suitable for the cultivation of the aquatic crops. Vegetable crops were irrigated during dry periods with overhead water emitted from perforated pipes. Crop yields were recorded for bordered areas from individual rows. Plant protection and other crop management practices followed AVRDC recommendations.
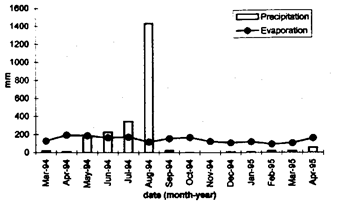
Fig. 1. Monthly cumulated
precipitation and evaporation in 1994/95 in AVRDC, Taiwan
Soil moisture tension measurement
Soil moisture tension was measured using vacuum gauge tensiometers installed within crop rows in one flat bed (one row), one 2.0-m-wide high bed (two rows), and two 3.0-m-wide high beds (three rows each) with two replications. Installation depth was 15 and 45 cm. Readings were taken at approximately two-day intervals from transplanting or seedling emergence until harvest of each crop. During the cultivation of each crop there were, however, periods when no readings were taken, partly due to inaccessible wet field conditions.
Calculation of soil moisture stress
To interpret variable soil moisture relationships with crop yield, soil moisture tension in the root zone can be calculated as a double integral of measurements over time of crop cultivation period and over soil depth. Including a correction factor for unequally spread time intervals between readings, the ‘integrated soil moisture tension’ is (Taylor, 1952 a):
|
|
|
[kPa]
|
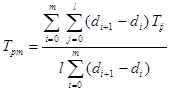
where: Tpm is the total moisture tension, i represents a single time, j represents a single depth, l is the total number of depths, m represents the total number of readings, d represents the day of the year when a reading was made, (di+1-di) is the time interval in days between successive readings, and Tij is the moisture tension at a single time and a single depth. In both equations the reference point is zero tension.
If an index of moisture stress should include stress caused by overwet soil conditions, the reference point of zero stress should be set to a value higher than zero tension. Stress can then be measured as the sum of the absolute value of the deviates from the optimum soil moisture tension. ‘Mean integrated soil moisture tension’ for time and soil depth then gives:
|
|
|
[kPa]
|
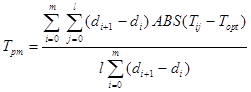
where: Topt is the optimum soil moisture tension. In this study mean integrated soil moisture tension was calculated for a single soil depth (15 cm), and for two depths (15 and 45 cm). Optimum soil moisture tension was defined for each crop where the relationship of crop yield with mean integrated soil moisture tension gave the best fit in the regression analysis.
Root-length-density measurement
Root length was measured in one flat bed and one 3.0-m-wide high bed shortly before final crop harvest by use of the ‘gridline intersect method’ introduced by Newman (1966). Soil was sampled with a 2.0-cm-diameter punch tube to a depth of 60 cm in distances of 20 cm from the edge towards the center of the beds with two replications. The soil column was cut into sections 10 cm long and the roots separated by carefully washing the soil through a fine (0.15 mm) sieve. The roots were spread out uniformly in a petri dish and put upon a grid of lines with an interline distance of 1.27 cm. This way, total root length in centimeter equals the number of counted root/gridline intersects (Giovannetti and Mosse, 1980). Through rearranging the roots in the petri dish, three readings were made for each sample. Root length density (cm/cm3) was calculated by dividing the mean of root length readings (cm) by the volume of the soil sample (cm3). Data from the topmost 10 cm of soil were excluded due to too many roots of weeds.
Results
Soil moisture tension
During the vegetable soybean crop in spring 1994 on average over the season soil moisture tension values were less and amplitude smaller on the flat bed compared to all positions within the high bed (Fig. 2 a, left). Differences between positions in the high bed were more conspicuous in the succeeding Chinese cabbage crop with the soil moisture in outer crop rows (40 and 80 cm from edge) reaching higher absolute tension than in the central row (Fig. 2 a, right). During chili, soil moisture approached low tensions from end of July until the middle of September particularly on flat beds and in the crop row close to the center of the 3.0-m-wide high bed (Fig. 3 a, left). During autumn and winter 1994 this changed to a periodic pattern of drying and re-wetting typical for fully irrigated field conditions. Soil moisture tension in flat beds averaged lower values with smaller amplitude than on high beds in which higher averages and greater deviations were recorded towards their centers. During cultivation of carrot in early 1995, soil moisture tension was greater on flat than on high beds (Fig. 3 a, right) throughout the cultivation period.
Mean integrated soil moisture tension
Calculated optimum soil moisture tension (Topt, Table 1) varied from a low level in carrot to higher tensions in crops of Chinese cabbage, vegetable soybean, and chili. Soil moisture tension below this reference point was considered as soil moisture stress (‘flooding’).
Table 1. Optimum soil moisture tension (Topt) for calculating mean integrated soil moisture tension and exponential regression (y = a × e(b×x); n = 9) of net yields (kg/m2) on mean integrated soil moisture tension (kPa) calculated over one soil depth (15 cm) and two soil depths (15 and 45 cm)
|
Mean integrated soil moisture tension |
Topt |
Regression analysis1 |
||
|
a |
b |
r2 |
||
|
Vegetable soybean |
|
|
|
|
|
15 cm depth |
23 |
1.73n.s. |
-0.026* |
0.42* |
|
15 and 45 cm depth |
11 |
1.56 n.s. |
-0.016* |
0.39* |
|
Chinese cabbage |
|
|
|
|
|
15 cm depth |
17 |
7.89 n.s. |
-0.746* |
0.74* |
|
15 and 45 cm depth |
16 |
13.28* |
-0.772 n.s.. |
0.65 n.s. |
|
Chili |
|
|
|
|
|
15 cm depth |
25 |
17.05 n.s. |
-0.308* |
0.63* |
|
15 and 45 cm depth |
18 |
1.65 n.s. |
-0.177 n.s.. |
0.42 n.s. |
|
Carrot |
|
|
|
|
|
15 cm depth |
8 |
3.42 n.s. |
-0.024 n.s.. |
0.19 n.s. |
|
15 and 45 cm depth |
8 |
3.28 n.s. |
-0.023 n.s. |
0.09 n.s. |
n.s.: not significant, *: significant at P = 0.05
Since vegetable soybean was particularly affected by stress caused by overdry soil conditions on high beds, stress curves did differ greatly between flat and high beds (Fig. 2 b, left). During cultivation of Chinese cabbage, moisture stress on high beds increased from the center towards the edge except for the first two weeks after transplanting when soil moisture tension was low (Fig. 2 b, right). Soil flooded conditions set in soon after transplanting chili in late July. Development of soil flooding was clearly reflected in the stress curves (Fig. 3 b, left). In this phase, stress indices were greatest for the flat bed closely followed by the innermost row on high beds. Integrated soil moisture stress declined more rapidly in the center of the high bed when soil moisture tension increased thereafter, approaching the tension in the outer rows. The influence of increasing soil moisture tension on moisture stress in the transition from wet season to dry season was reflected in a higher rate of increase in mean integrated soil moisture tension towards the inside row of the 3.0-m-wide high bed. Overdry soil conditions prevailed during the carrot crop in early 1995, similar to the vegetable soybean crop in spring 1994, although less pronounced. Mean integrated soil moisture tension was higher in flat beds compared to high beds throughout cultivation (Fig. 3 b, right). Final stress indicators for 3.0-m-wide high beds were smaller close to the edge and to the center.
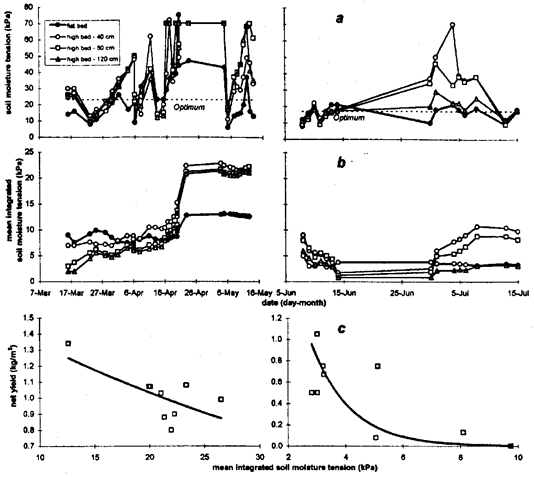
Fig. 2. Soil moisture tension (a), mean integrated soil moisture tension (b) at 15 cm soil depth, and the exponential relationship between mean integrated soil moisture tension and net yield (c) for (left) vegetable soybean and (right) Chinese cabbage in two cultivation systems and different crop row positions. The dotted horizontal line in (a) indicates the 'optimum soil moisture tension'.
Regression of yield on mean integrated soil moisture tension
Yields of vegetables during the dry season (vegetable soybean and carrot) were more linearly related to the mean integrated soil moisture tension (Fig. 2 and 3 c), whereas the relationship between yield and soil moisture stress was exponential for vegetables (Chinese cabbage and chili) during the rainy season. The exponential regressions were significant when soil moisture stress was measured at 15-cm depth and were not improved upon by inclusion of data from the lower (45-cm) soil depth (Table 1). However, the equations were not suitable to estimate maximum yields of crops when no moisture stress occurred since the intercept was usually not significant. For carrot there was no clear relationship between yield and integrated moisture tension.
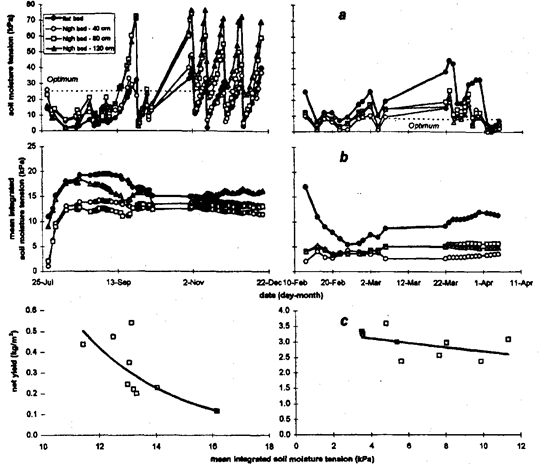
Fig. 3. Soil moisture tension (a), mean integrated soil moisture tension (b) at 15 cm soil depth, and the exponential relationship between mean integrated moisture tension and net yield (c) for (left) chili and (right) carrot in two cultivation systems and different crop row positions. The dotted horizontal line in (a) indicates the 'optimum soil moisture tension'.
Gradients of soil moisture tension within high beds
The gradient in soil moisture tension between 15 and 45-cm soil depth increased with increasing absolute tension at the shallow depth. When the soil was rewetted during the irrigation cycles in November and December 1994 (Fig. 4), this gradient became negative, indicating that the topmost soil layer was wetter than the deeper layer. The soil was saturated throughout the profile from end of July until early September. This was reflected in low absolute tension at 15-cm depth and a small gradient between 15 and 45-cm soil depth. Under these conditions water moved from the inside towards the outside of the high bed, indicated by a positive horizontal gradient. With increasing soil moisture tension and increasing gradient between shallow and deeper soil layer, water flow was increasingly directed towards inside of the high bed.
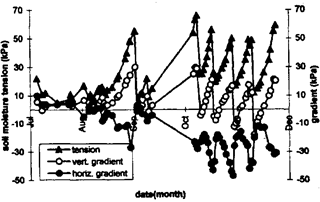
Fig. 4. Soil moisture tension at 15 cm soil depth, vertical gradient of moisture tension (15-to-45 cm depth), and horizontal gradient of moisture tension (40 -to-120 cm distance from edge) for chili (a positive gradient indicates upward water flow and outward water flow).
Root-length-density distribution
Root-length density was typically restricted to the top 50-cm soil depth (Fig. 5 a to d). Root density in the whole profile was greater in high beds (right) for all vegetables. Fewer roots were found above 20-cm depth, but roots elongated more profusely in the 20 to 40-cm soil layer.
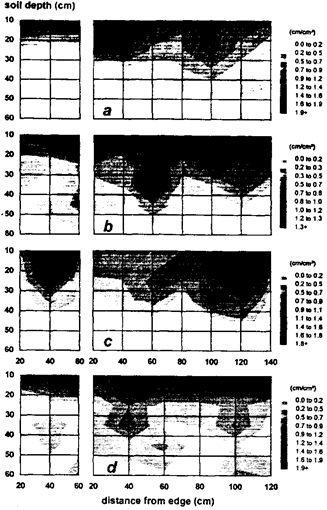
Fig. 5. Root-length-density distribution of (a) vegetable soybean, (b) Chinese cabbage, (c) Chili, and (d) carrot in the (left) 1.5-m-wide flat bed and (right) 3.0-m-wide high bed.
Discussion
Soil moisture stress and optimum soil moisture tension
One limitation in the determination of soil moisture stress may have been the use of vacuum gauge tensiometers. These tensiometers function only over a limited part of the plant available water range (Hanks and Ashcroft, 1980). Some authors (Gardner, 1960; Gingrich and Russell, 1957) have indicated that plant growth is only affected at much higher soil moisture tension, but for snapbean yield was found to decrease very sharply with increasing tension in the low-tension range (Millar and Gardner, 1972).
Gardner (1964) stated that once root distribution in the soil profile is known, measurement of soil moisture tension at a single appropriate depth was sufficient. Since roots were restricted to the upper 40-cm soil depth, inclusion of soil moisture tension at 45-cm soil depth did not improve the estimation of yield as a function of soil moisture stress. In retrospect installation of tensiometers at a depth of 20 cm would be sufficient under our conditions.
Evaporation largely exceeded precipitation during vegetable soybean in the dry spring season (Fig. 1), and water stress was primarily related to stresses caused by overdry soil conditions. Chinese cabbage and chili grown during the summer rainy season and subsequent early dry season were affected by stresses caused by both, excess and deficit soil water. Under these conditions, the relationship between soil moisture stress and yield was clearly exponential. The high values of parameter b in the equations (Table 1) indicate that extreme values of soil moisture tension exerted an exaggerated effect on crop growth. Since this was true only in the rainy season, overwet soil conditions indicated by low moisture tensions explained the greater part of variations in yield. The insignificant estimates of maximum crop yield when no soil moisture stress occurred (parameter a) were partly an extrapolation problem since no real soil moisture treatments were imposed and the number of observations was limited. The study accounted for crop-specific sensitivities to water stress (Topt) but not for the fact that these sensitivities may also vary with stage of crop growth (Hiler et al., 1972). The influence of soil moisture stress on carrot yield was insignificant presumably because soil moisture tension was held near-optimum throughout the cultivation period so that soil moisture was not a growth-limiting factor. Additional effects of nitrogen availability (Kleinhenz et al. 1996) were not considered here but might have played a role.
Optimum soil moisture tensions calculated for individual crops showed increasing tolerance to overwet soil conditions in the order: chili, vegetable soybean, Chinese cabbage, and carrot. Chili is particularly sensitive to soil inundation. Flooding induced decline in photosynthesis, resulting in reduction in leaf area, plant weight, and dry matter accumulation (AVRDC, 1989). The ability of grain soybeans to acclimatize to saturated soils in seasonally waterlogged tropical lowland areas was assumed to have developed during the long period of domestication in rice-based Asian agriculture (Lawn, 1985). However, prolonged flooding may significantly reduce soybean growth (Sallam and Scott, 1987). This was found to be due to its sensitivity to low oxygen concentrations even when soil matric potential was maintained close to optimum (Sojka, 1985). Soybean lines for vegetable consumption are particularly sensitive to unbalanced water supply. Flooding in Chinese cabbage may not only cause a reduction in plant growth, but can also lead to a complete destruction of the root system (AVRDC, 1980). However, sufficient soil water must be present to compensate for transpiration loss and to facilitate calcium uptake (Yingjajaval, 1990), a prerequisite to avoid the physiological disorder of ‘tipburn’ (Imai, 1987). Topt for Chinese cabbage was found to be lower than for chili and vegetable soybean in this study, pointing out the importance of well balanced, yet sufficient soil moisture. Although wet soil conditions may reduce market quality in carrots, high and particularly steady water supply is a prerequisite for high yields (Krug, 1991).
Soil moisture stress, root growth and yield
Although differences in root growth characteristics were anticipated, they varied not much among the four vegetable species. Even though plant species have individual root growth characteristics, these can be substantially modified by environmental conditions: cultivated plants subjected to drought often develop deep, profusely branched root systems to absorb water and nutrients from a large volume of soil. However, when grown with irrigation and fertilization, smaller root systems may be sufficient (Kramer, 1983). Greater root growth under those conditions may only indicate partitioning of greater energy to the root system and not an increase in water and nutrient uptake (Devitt, 1989). In hydroponics, saturated soil culture, areas with high water tables, or under high irrigation rates, roots accumulate close to the soil surface (Protopapas and Bras, 1987). Roots of vegetables in this experiment typically accumulated above 40-cm soil depth. This must be viewed in the context of transformation of paddy rice fields to upland crop production (Kleinhenz et al. 1995).
When soil moisture was temporarily deficient during cultivation of vegetable soybean, roots elongated more profoundly to deeper soil layers in high beds. Yields were, however, lower than on high beds, suggesting that too much photosynthate was diverted into root growth at the expense of shoot growth and yield. Other reports (e.g., Heatherly, 1980) show that more root was required to support soybean shoot growth and development when grown in dry soil.
In more flood-prone flat beds, root systems of vegetables were typically restricted to the uppermost soil layer during the rainy season. Flooding may lead to the death of deeper roots and often the proliferation of adventitious and surface roots. This may expose them to more favorable chemical and physical conditions (Jones et al., 1991), but can make them more sensitive to subsequent drought (Jackson and Drew, 1984). Formation of lysigenous aerenchyma and adventitious root close to the soil surface (AVRDC, 1993) are important plant responses in chili for survival under flooded conditions. Yields on flat beds were, however, much lower than on high beds, indicating that adventitious rooting may have helped chili to recover from flooding, but that these roots may have only incompletely replaced the functions of the original roots (Jackson and Drew, 1984).
Implications for cultivation systems
High bed cultivation appeared to be a good solution for production of flood-prone vegetables during the wet season in the rice-based tropical environment investigated in this study. This is based upon improved hydraulic conditions in high beds. In tropical lowlands, water tables are frequently close to the surface during the rainy season. Although the surface soil may be distinctively drier than the water-logged soil beneath, water supply through rainfall will quickly exceed the soil’s limited rate of absorption. Under such conditions, decrease in vertical infiltration results from the decrease in moisture-potential gradient (Hillel, 1980). If surface runoff is limited, flat planting beds will become entirely water-logged then. Furrows between high beds act as a drain into which excessive soil water flows along a horizontally directed hydraulic gradient between inside and outside of the bed. Figure 4 revealed that low soil moisture tension and a small vertical gradient are both a prerequisite for horizontal drainage. During the rainy season a sink, the flooded furrows acted as a source to supply high beds with water during the dry season. Crop demand and evaporation deplete soil water in the surface layer. When moisture tension in this layer and the gradient between topsoil and subsoil was high, water flow from the edge towards center was maximal.
Since high beds can be constructed with different dimensions, adjustment of height and width is of primary interest. Height of beds depends primarily on width of initial construction since the latter determines the amount of soil material available when width of furrows is fixed. It was shown that the inside of wider high beds is more rapidly inundated as heavy rainfall prevails, that excessive soil water will be more quickly removed soon after (Fig. 3 b, left) and that moisture will become more rapidly deficient in periods of limited rainfall if soil moisture is not closely controlled by irrigation. If space allocation to high bed (vegetable) cultivation area is to be maximized for a given unit of land (as in intensive peri-urban horticulture) and furrow width is fixed, then high beds should be as wide as practical. But, if soil inundation through continuous rainfall usually proceeds for prolonged times and if efficient irrigation facilities are not available to overcome deficient soil water during periods of limited rainfall, narrower beds are called for. However, irrigation proved to be crucial for crop production during dry periods since the low height of standing water suitable for aquatic crop production in the furrows could only supply a part of crop water needs.
Acknowledgments
We thank the German Federal Ministry of Economic Cooperation (BMZ) for funding and AVRDC for its support in the implementation of this project.
References
AVRDC 1980. Progress Report for 1979. Asian Vegetable Research and Development Center, Shanhua, Taiwan.
AVRDC 1989. Progress Report for 1988. Asian Vegetable Research and Development Center, Shanhua, Taiwan.
AVRDC 1993. Progress Report for 1992. Asian Vegetable Research and Development Center, Shanhua, Taiwan.
Bathke, G. R., Cassel, D. K., Hargrove, W. L. and Porter, P. M. (1992). Modification of soil physical properties and root growth response. Soil Sci. 154: 316-329.
Brady, N. C. 1990. The Nature and Properties of Soils. Macmillan, New York.
Callebaut, F., Gabriels, D., Minjauw, W. and De Boodt, M. 1982. Redox potential, oxygen diffusion rate, and soil gas composition in relation to water table level in two soils. Soil Sci. 134, 149-156.
Cassell, D. K. and Klute, A. (1986). Water potential: tensiometry. In: Methods of Soil Analysis, Part 1, Physical and Mineralogical Methods, Second Edition (Ed. A. Klute), pp. 563-596. ASA, SSSA, Madison, WI.
Chiu, C. C. 1987. Evolution of farming systems in Taiwan. ASPAC Extension Bulletin 265. FFTC, Taipei, Taiwan.
Devitt, D. A. 1989. Bermudagrass response to leaching fraction, irrigation salinity, and soil types. Agron. J. 81, 893-901.
Gardner, W. R. 1960. Dynamic aspects of water availability to plants. Soil Sci. 89, 63-73.
Gardner, W. R. 1964. Relation of root distribution to water uptake and availability. Agron. J. 56, 41-45.
Gingrich, J. R. and Russell, M. B. 1957. A comparison of soil moisture tension and osmotic stress on root growth. Soil Sci. 84, 185-194.
Giovannetti, M. and Mosse, B. 1980. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phyt. 84, 489-500.
Hanks, R. J. 1991. Soil evaporation and transpiration. In: Modelling Plant and Soil Systems (Eds J. Hanks and J. T. Ritchie), pp. 245-272. ASA, CSSA and SSSA, Madison, WI.
Hanks, R. K. and Ashcroft, G. L. 1980. Applied Soil Physics. Springer Verlag, Berlin.
Heatherly, L. G. 1980. Growth of soybeans at different soil matric potentials. Soil Sci. 130, 331-335.
Hiler, E. A., van Bavel, C. H. M., Hossain, M. M. and Jordan, W. R. 1972. Sensitivity of southern peas to plant water deficit at three growth stages. Agro. J. 64, 60-64.
Hillel, D. 1980. Applications of soil physics. Academic Press, New York.
Imai, H. 1987. NH4-N toxicity and calcium deficiency in tipburn and internal rot in Chinese cabbage. Technical Bulletin No. 105. ASPAC Food and Fertilizer Technology Center, Taipei, Taiwan.
Jackson, M. B. and Drew, M. C. 1984. Effects of flooding on growth and metabolism of herbaceous plants. In Flooding and Plant Growth (Ed. T. T. Kozlowski), pp. 47-128. Academic Press, New York.
Jones, C., Bland, W. L., Ritchie, J. T. and Williams, J. R. 1991. Simulation of root growth. In Modelling Plant and Soil Systems (Eds J. Hanks and J. T. Ritchie), pp. 91-124. ASA, CSSA and SSSA, Madison, WI.
Kieft, J. 1994. Between plants and business. Farming styles in the Bangkok vegetable industry. M. Sc. thesis, Agricultural University Wageningen.
Kleinhenz, V., Schnitzler, W. H. and Midmore, D. J. 1995. High bed systems for off-season vegetable production in the Tropics and Subtropics. Entwicklung und Ländlicher Raum 4, 26-28.
Kleinhenz, V., Schnitzler, W. H. and Midmore, D. J. 1996. Managing nitrogen fertilization for year-round vegetable production in paddy rice fields. Gartenbauwissenschaft 61, 25-32.
Kramer, P. J. 1983. Water Relations of Plants. Academic Press, New York.
Krug, H. 1991. Gemüseproduktion. Verlag Paul Parey, Berlin.
Lawn, R. J. 1985. Saturated soil culture - expanding the adaptation of soybeans. ACIAR Foodlegume Newsletter 3, 2-3.
Midmore, D. J., Roan, Y. C. and Wu, M. H. 1995. Management practices to improve lowland subtropical summer tomato production: yields, economic returns, and risk. Expl. Agric. (in press).
Millar, A. A. and Gardner, W. R. 1972. Effect of the soil and plant water potentials on the dry matter production of snapbeans. Agro. J. 64, 559-562.
Newman, E. I. 1966. A method of estimating the total length of root in a sample. J. App. Ecol. 3, 139-145.
Plucknett, D. L. and Beemer, H. L. (Eds) 1981. Vegetable Farming Systems in China. Westview Press, Boulder.
Ponnamperuma, F. N. 1984. Effects of flooding on soils. In: Flooding and Plant Growth (Ed. T. T. Kozlowski), pp. 10-46. Academic Press, New York.
Protopapas, A. L. and Bras, R. L. 1987. A model for water uptake and development of root systems. Soil Sci. 144, 352-366.
Rawitz, E. and Hillel, D. I. 1969. Comparison of indices relating plant response to soil moisture status. Agro. J. 61, 231-235.
Richter, J. 1987. The Soil as a Reactor: Modelling Processes in the Soil. Catena Verlag, Cremlingen.
Sallam, A. and Scott, H. D. 1987. Effects of prolonged flooding on soybeans during early vegetative growth. Soil Sci. 144, 61-66.
Sjahri, S. 1975. Crop intensification in rainfed paddy areas in Indonesia. In: Proceedings of the Cropping Systems Workshop March 18-20, 1975, pp. 57-68. IRRI, Los Baños.
Sojka, R. E. 1985. Soil oxygen effects on two determinate soybean isolines. Soil Sci. 140, 333-343.
Taylor, S. A. 1952 a. Estimating the integrated soil moisture tension in the root zone of growing crops. Soil Sci. 73, 331-339.
Taylor, S. A. 1952 b. Use of mean soil moisture tension to evaluate the effect of soil moisture on crop yields. Soil Sci. 74, 217-226.
Wadleigh, C. H. 1946. The integrated soil moisture stress upon a root system in a large container of saline soil. Soil Sci. 61, 225-238.
Yingjajaval, S. 1990. Effect of water and nitrogen on Chinese cabbage. Kasetsart J. (Nat. Sci.) 24, 244-252.